Prírodou inšpirované algoritmy
študijné materiály pre projekt mobilnej triedy umelej inteligencie

Rastlina a okolie
Exogénna a endogénna regulácia
Difúzne ohraničené zhlukovanie a cellulárne automaty poskytujú modely s exogénnym mechanizmom tvarovania vetviacich štruktúr. V takom prípade prvky narastajúcej štruktúry komunikujú cez okolitý priestor. V protiklade k tomu, L-systémy predstavujú endogénne riadený mechanizmus, ktorý sa spolieha na informačný tok vo vnútri štruktúry. Prenášanie informácia dedením a vzájomná interakcia medzi susednými modulmi sú endogénneho riadenia. V prírode sú endogénne a exogénne riadenie často kombinované. Napríklad, vývoj stromu je urobený pomocou geneticky riadeného formovamia kvázi buniek, prúdenia vody, výživných látok a fytohormónov cez vetviace štruktúry a cez odozvu na faktory prostredia ako napr. nedostatok svetla alebo hustotu vetiev.
Enviromentálne L-systémy
Enviromentálne L-systémy predstavujú jedno z priblížení sa vytvoreniu úplného modelu zjednocujúceho endogénne a exogénne javy (Prusinkiewicz, James & Mech,1994). V tomto prípade sa predpokladaný vývoj uskutočňuje v priestore charakterizovanom číselne alebo skalárne. Moduly rastúcej rastliny testujú hodnoty poľa v oblasti na ktorú majú nárok.
Napríklad animácia ukazuje proces v enviromentálnom L-systéme simulujúcom reakciu na 2D ohraničenie vetviacich štruktúr orezaním. Rozvíjaná štruktúra je obmedzená na štvorcovú oblasť a vrcholky testujú či sú vnútri alebo vonku tejto oblasti. Počas úvodnej fázy vývoja, vrchol hlavnej stonky ju predlžuje a vytvára postupnosť nevyužitých zárodkov. Po prekročení hranice štvorca je vrchol odrezaný a signál aktivujúci najbližší spiaci zárodok je poslaný basipetálne. Aktívny zárodok vytvorí postrannú vetvu, ktorá rastie tým istým spôsobom ako počiatočná štruktúra (traumatic reiteration). Po prekročení hraničného štvorca je nový vrchol odrezaný a signál aktivujúci zárodok znovu aktivovaný. Konečná štruktúra je výsledkom viacnásobného opakovania tohto procesu. 3D rozšírenie tohto modelu je ukázané v animácií (bez listov) a v animácií(s listami). V oboch prípadoch, niektoré novovytvorené zárodky vytvárajú nové štruktúry podobné zasadeným stromom.




Orezávanie vetiev vytvára ornamentálne útvary nazývané topiary. Napríklad obrázok a obrázok ukazuje objekt, kde ohraničenie je definované ako cylinder. Obrázok ukazuje dvojicu stromov orezaných ako špirála. Obrázok spája tieto a ďalšie stromy do umelého obrazu topiary záhrady, inšpirácia je z Levens Hall garden v Anglicku (Coats,1963). obrázok reprezentuje ďalší komplex ohraničení podobný dinosauru. Jeho povrch je vytvorený tak, že pokrýva kostru z línii a elipsoíd. Obrázok ukazuje výsledný útvar ako strom s listami.


Interakcia medzi rastlinami a ich okolím
Topiary sú príkladom modelov rastlín, v ktorých má vplyv prostredie. V prírode, interakcia medzi rastlinou a jej okolím má často zložitejší charakter, prostredie pôsobí na rastlinu a tá opačne pôsobí na prostredie. Tento obojsmerný tok informácií môžeme chápať ako spätnoväzobnú slučku ukázanú na obrázku dole. Simulácia rastlinnej interakcie s okolím môže byt vykonaná vnútri všeobecného rámca open L-systémov (Mech & Prusinkiewicz,1996). Predpokladá sa vyvíjanie v priestore charakterizovanom skalárnym alebo vektorovým polom. Moduly rastúcej rastliny môžu testovať hodnoty tohto poľa ako body podľa dôležitosti a posielať hodnoty, ktoré ovplyvnia pole v určitej oblasti.
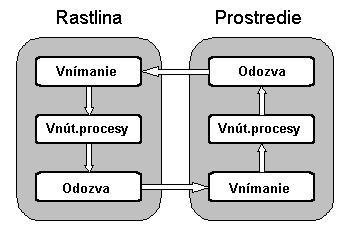
Informačný tok medzi rastlinou a prostredím
Animácia ukazuje L-systém tvoriaci jeden z prvých modelov rastlinám podobných vetviacich štruktúr, navrhnutý v roku 1967 Cohenom (Cohen ,1967). Tento model má jemnú lokálnu hustotu rastúcej štruktúry. Gradient funkcie hustoty sa používa na vybratie najmenšej plochy na ďalšie rozvíjanie každej vetvy. Vo veľmi hustej oblasti sa rast zastavuje úplne. Týmto spôsobom je výsledný obraz určený prostredím sprostredkovanou interakciou medzi vetvami.
Animácia ukazuje rozvíjanie dvoch planárnych vetiev, ktoré bojujú o priestor. Základný model bol popísaný na základe pozorovania tropického stromu Terminalia catappa (indický mandľovník) a navrhli ho Honda, Tomlinson a Fisher (Honda et al.,1981). Kruhy reprezentujú skupiny listov lokalizované v uzloch. Koncový bod každej vetvy alebo vrcholu produkuje nové vetvy, len ak by mohli padnúť do existujúcej skupiny. Táto interakcia limituje rozsah vetiev a prispôsobuje formu každej vrstvy v súvislosti s jej susedom.
Animácia ukazuje pohľad zvrchu na oblasť zeme s rôznou intenzitou osvetlenia. Hypotetická klonovaná rastlina (inšpirovaná ďatelinou) sa množí prostredníctvom horizontálnych stoniek (spacers) ktoré pripájajú jednotlivé rastliny (ramets). Staré spacers a ramets odumierajú. Klon využíva vysokú intenzitu svetla pri zvyšujúcej sa hustote vetiev a klesajúcej dĺžke stoniek. Kolíziám je zabránené ako v predošlej simulácii. Po kolonizácii najobľúbenejšieho ľavého dolného miesta rastlina robí inváziu do pravej hornej časti. Svetelné podmienky v tejto časti však nie sú dostatočné pre plynulé roztiahnutie rastliny. Kolónia sa stráca, až kým je oblasť dosiahnutá znova novou vlnou množenia. Dynamika množenia odzrkadľuje adaptáciu rastliny na jej prostredie.
Animácia ukazuje 2D model koreňa hľadajúceho vodu v pôde počas jeho rastu. Počiatočná distribúcia vody je vopred daná v tvare S zóny s vysokou koncentráciou indikovanou svetlou farbou. Narastanie nakláňa hlavný koreň. Malé korienky absorbujú vodu rozptýlenú v pôde. Ubúdajúca koncentrácia vody je indikovaná tmavými plochami ktoré sa objavujú okolo koreňového systému. V oblastiach s nedostatkom vody sa koncentrácia korienkov znižuje. ešte predtým než dosiahnu svoju potencionálnu dĺžku.
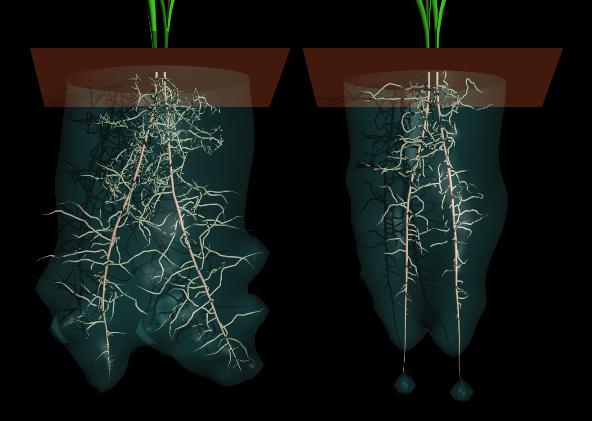
Animácia ponúka 3D rozšírenie predchádzajúceho modelu, založené na práci Clausnitzera and Hopmansa (Clausnitzer & Hopmans,1994). Koncentrácia vody je vizualizovaná prostredníctvom polopriehľadného povrchu okolo koreňov. Ako vyplýva zo súťaženia pri hľadaní vody hlavné korene rastú vzájomne od seba. Ak menšie korienky rastú pomaly, oblasť účinku každého koreňového systému je menšia a korene rastú bližšie pri sebe. Tento efekt je ilustrovaný na obrázku.


Obrázok a obrázok ukazuje modely gaštana inšpirované prácou Chiba (Chiba et al.,1994) a Takenaka (Takenaka,1994). Konáre stromu si konkurujú pri prijímaní svetla z oblohy. Skupiny listov robia tieň nižším vetvám. Vrchol v tieni neprodukuje nové vetvy. Existujúce vetvy ktoré neprijímajú svetlo odumierajú a sú zhodené zo stromu. Takýmto spôsobom boj o svetlo riadi hustota vetiev v korune stromu.
V animácií dva geneticky identické stromy bojujú o svetlo. Ich následné oddelenie po ukončení vývoja odhaľuje ich prispôsobenie susednému stromu. Podobnú adaptáciu pozorujeme na ihličnatých stromoch na obrázku a obrázku.


Obrázok ďalej ilustruje dopad boja o svetlo. Stromy na okraji majú asymetricky rozvinuté koruny a chýbajú im niektoré spodné vetvy. Stromy medzi nimi nemajú spodné vetvy pretože nemajú dostatok svetla. V drevárskom priemysle je chýbanie spodných vetiev veľmi žiadaný stav, keďže sa tým redukujú uzly v kmeni a čas potrebný na očistenie kmeňa. Takáto simulácia pomáha pri určení optimálnych vzdialeností pri vysádzaní stromov.
