Prírodou inšpirované algoritmy
študijné materiály pre projekt mobilnej triedy umelej inteligencie

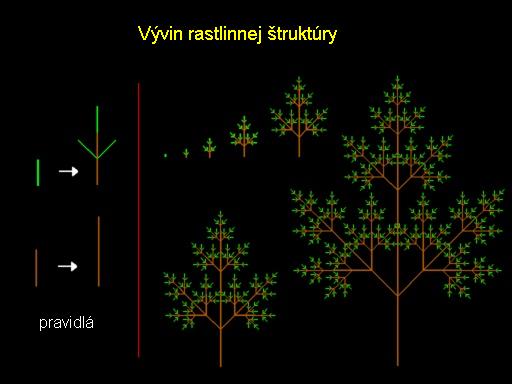
Princípy v modelovaní rastlín
Vetviace vzory rastlín.
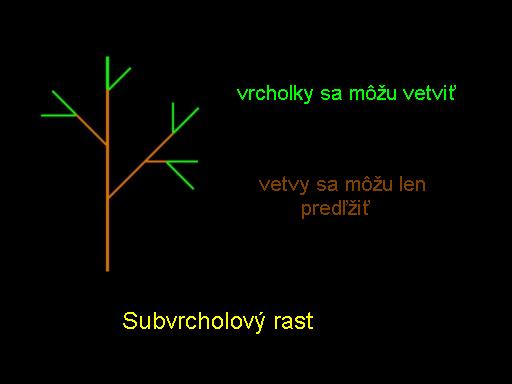
Napriek svojej jednoduchosti model rastu listov ukázaný na obrázku vhodne ukazuje jeden zo základných princípov vývoja rastlín. Ide o princíp subvrcholového rastu t.j. že iba vrchol (apex) môže vytvárať nové vetvy. Stonka (časť medzi dvomi uzlami) môže byť predĺžená, ale nemôže začínať vetvy. Tieto pravidlá sú schematicky ilustrované na obrázku.
{kind=link}
Monopodálne vetvenie
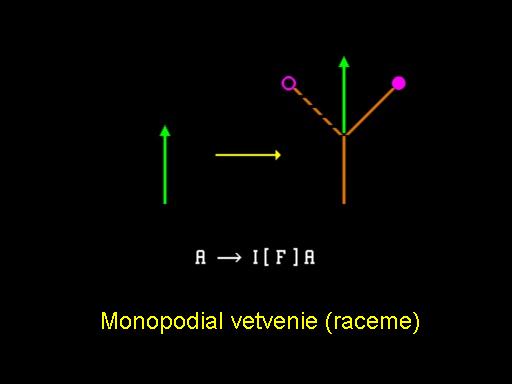
Rozmanitosť štruktúr môže byť vytvorená cez subvrcholový rast v závislosti na tom, ktorý vrchol zostane aktívny a iniciuje vetvenie a ktorý nie. V súvislosti s výskumom Lindenmayera a iných biológov používajúcich L-systémy, táto časť je zameraná na modelovanie a simuláciu rastu súkvetí (inflorescences) rastlín. Podrobnejší popis je v (Prusinkiewicz & Lindenmayer,1990).

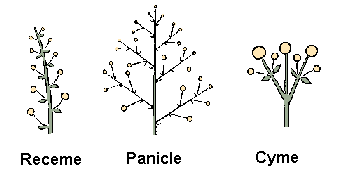
Najjednoduchší prípad nastane, keď iba vrchol hlavnej osi narastajúcej rastliny tvorí nové vetvy, ako ukazuje obrázok. Na týchto postranných vetvách sa vytvoria púčiky a neskôr kvety, ale netvoria ďalšie vetvy. Výsledná štruktúra sa nazýva monopodiálna vetviaca štruktúra a príslušné vetvenie je nazývané strapec (raceme). Obrázok ilustruje vývoj bežnej buriny Capsella bursa-pastoris (pastierskej kapsičky) sledujúc tento princíp. Campanula ranunculoides (zvonček iskerníkovitý), ukázaný v animácií, sa vyvíja tým istým spôsobom. Dodatočné pohľady na tento vývoj sú v animácií , kde sú spojené modely niekoľkých rastlín na jednej scéne a v animácií, kde je prezentovaný pohľad na kvet.
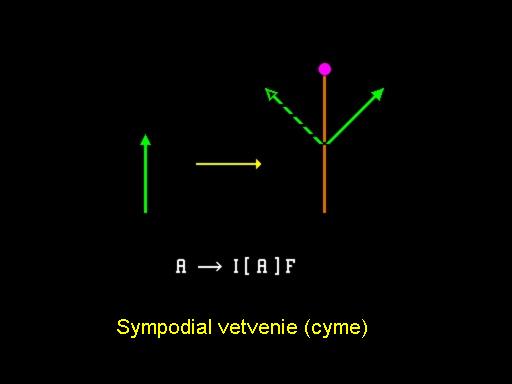
Sympodiálne vetvenia
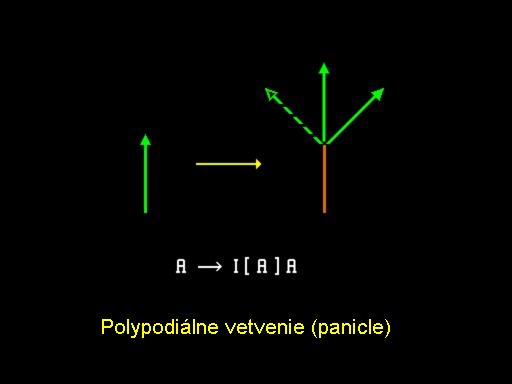
Polydiálne vetvenia
Typy súkvetí (inflorescences)
Obrázok ukazuje spôsob vetvenia, ktorý je vnímaný ako protiklad k predošlému. V tomto prípade nazývanom sympodiálne vetvenie, vrchol produkuje púčiky kvetov, ktoré ukončujú vývoj v danej osi a naviac produkuje jeden alebo viac aktívnych postranných vrcholov. Tento proces sa rekurzívne opakuje s hlavným tlakom vývoja vždy idúcim do postranných vetiev. Výsledný typ vetvenia sa nazýva vidlica (cyme). Vývoj Lychnis coronaria (kukučky vencovej) ukázaný v animácií a animácia, poskytuje dobrý príklad popísaného procesu. Animácia prezentuje pohľad na vývoj jedného kvetu. Ďalší typ vetvenia, nazývaný polypodiálne vetvenie, je uvedený schématicky na obrázku. V tomto prípade aj vrchol v hlavnej línii aj postranné vrcholy tvoria nové vetvy až kým sa v istom čase zapne tzv. vývojový prepínac a vrcholky sa transformujú na púčiky kvetov. Výsledná štruktúra sa nazýva metlina (panicle) a môže mať rôzne formy v závislosti na geometrických parametroch. Napríklad obrázok ukazuje tzv. krížové vetviace obrazce, kde páry vedľajších vetiev sú usporiadané vo vzájomne kolmých rovinách. Ukončením každej z týchto vetiev kvetom môžeme modelovať štruktúru orgovánu, ako ukazuje obrázok.
{kind=link}

Krížové vetvenia

Model orgovánu
Signály v rastlinách
Mohli by sme sa domnievať, že teraz sme schopní modelovať všetky možné typy rastlín, s ohľadom na možné vetviace štruktúry. Nie je to celkom pravda. Jeden problém sa týka sekvencie kvitnutia. Napríklad, v divej mrkve, ukázanej na obrázok , kvety sa najprv objavia na vrchole štruktúry a sféra kvitnutia postupuje smerom nadol. Zatiaľ čo rastlina sa vyvíja zdola nahor a nie od vrchu nadol, poradie kvitnutia je opačné ako poradie v ktorom boli púčiky formované. Mäta na obrázku ilustruje tento jav. Vrchná skupina kvetín bola vytvorená posledná, ale rozkvitla ako prvá.


Za účelom zachytenia takých procesov je potrebné vziať do úvahy rôzne formy toku informácii v rozvíjanej rastline. Pravidlá v L-systémoch opísané v predtým znázorňujú transfer informácie dedením (pôvod, rodokmeň), alebo z hlavného modulu do dcérskeho modulu. Tieto pravidlá sa vo formálnom jazyku nazývajú bezkontextové. V mnohých rastlinách vzájomné pôsobenie medzi existujúcimi časťami tiež hrá dôležitú úlohu. Takáto interakcia môže mať formu toku výživných látok, vody alebo hormónov medzi časťami vyvíjanej rastliny. Vzájomný vzťah medzi tokom informácií daným geneticky a vzájomným pôsobením je schematický ukázaný na obrázku. Bunka (modul) je rozdelená, produkuje dve nové bunky (moduly), ktoré sa neskôr tak isto rozdelia. Informácia môže byť prenášaná postupne od materského modulu k dcérskemu (diagonálne šípky) alebo pomocou vzájomnej interakcie (horizontálne šípky).
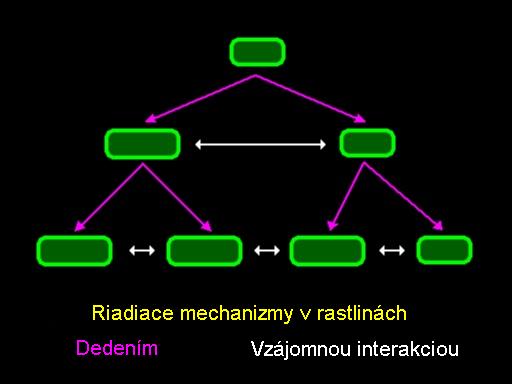
Riadiace mechanizmy v rastlinách
Šírenie signálu
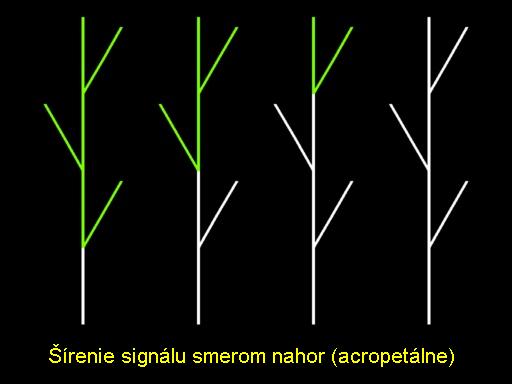
Interaktívne procesy môžu byť realizované kontextovými L-systémami. V takom prípade pravidlo je aplikované v čiastkovom module len ak tento modul má nejakých špecifických susedov. Napríklad, na obrázku zelený modul sa stáva biely vtedy a len vtedy ak je priamo pod ním biely modul. Takto cez kontextovú súvislosť je možné simulovať šírenie signálu.
Tok informácií v statickej štruktúre je v niečom zaujímavý, ale oveľa zaujímavejšie javy sa objavujú v rastúcich štruktúrach. Napríklad, vezmime do úvahy signál indukujúci kvitnutie, odpovedajúci hypotetickému hormónu florigén, ktorý je poslaný z dolnej časti rastliny smerom k vrcholu. Ak vrchol prijme tento signál, je transformovaný do kvetinového púčika a neskôr na kvet a samotný plod. V závislosti na pomere rýchlosti rastu hlavnej stonky a postranných vetiev v príslušnej úmernosti šírenia signálu, oblasť kvetín môže postupovať akropetálne (smerom od stredu k okrajom) ako ukazuje animácia, alebo basipetálne (smerom nadol) ako ukazuje animácia. Animácia a animácia ilustruje basipetálne poradie kvitnutia použitím schematického a realistického modelu púpavy. Keď vrchol hlavnej stonky bol transformovaný na kvetinový puk, kvety na dolných vetvách sú formované postupne podľa basipetálnej postupnosti. Animácia ukazuje vývoj jedného kvetu od púčika cez otvorenú kvetinu až po odkvitnutie.
Niektoré modely požadujú niekoľko signálov. To nastáva hlavne v prípadoch, keď sa postupnosť kvitnutia vyvíja smerom nadol a v niektorých častiach je rastlina rozvinutejšia v blízkosti vrcholu než v blízkosti spodnej časti. Príklad nám poskytuje rastlina ukázaná v animácií. Spočiatku vrchol hlavnej stonky produkuje postupne vedľajšie vetvy. V tejto etape ich ďalší rozvoj brzdí. V istom okamihu je florigén indukovaný a šírený smerom nahor. Ak dosiahne vrchol stonky, tento sa mení na púčik a neskôr na kvet. Krátko nato je iný signál poslaný z vrcholu a šírený nadol. Tento signál zvyšuje hodnotu v uzloch umožňujúcu postranným vetvám narastať. Ak je umožnený takýto proces zhora nadol, vrchná časť štruktúry môže byť viac rozvinutá než dolná časť. Tretí signál (nie je ukázaný v animácii) privádza florigen do postranných vetiev, tak že ich vrcholy sa môžu nakoniec transformovať na kvety. 3D obrázok tohto procesu je ukázaný na obrázku a animácií. Signály môžu byť použité tiež na simuláciu nejakých procesov mimo rastliny. Napríklad v animácií signál reprezentuje hmyz skúmajúci rastúcu rastlinu a požierajúci jej vrcholky. Vetvy, ktoré už nemajú vrcholky zvädnú. Podobné modely môžu byť konštruované pri predpoklade rôznych stratégií pre jeden alebo viacero druhov hmyzu (ktoré sa môžu vzájomne ovplyvňovať).
