Prírodou inšpirované algoritmy
študijné materiály pre projekt mobilnej triedy umelej inteligencie
|
|||||||||||||

Genetické regulačné siete
(Dellaert, 1995) vyzdvihol fakt, že výpočtová komplexnosť nízkoúrovňových simulácií môže spôsobiť evolúcii ťažkosti. Preto Dellaert a Beer navrhli ambiciózny a rozsiahly model vývinu, ktorý mal byť biologicky obhájiteľný a výpočtovo zvládnuteľný. Model sa zameriava na myšlienku Genetických regulačných sietí (GRN), čo sú siete interakcií medzi genómmi a ich prostredím, ktoré vedú k postupnosti zmien stavov vnútri každej bunky.
Dellaert a spol. implementovali ich GRN použitím boolovských funkcií nazvaných „operóny“ vnútri genómu. Pre ilustráciu zvážte nasledovné dva operóny
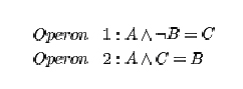
Prvá rovnica znamená, že ak proteín A je prítomný v bunkovej cytoplazme a proteín B nieje, potom sa produkuje proteín C a je pridaný do cytoplazmy. Prítomnosť proteínu A a C zabezpečuje druhý operón, ktorý vytvára B v cytoplazme po prvý krát. V tomto príklade, kde operón 1 iba produkuje proteín C len za neprítomnosti B, by bunka prestala produkovať proteín C. Reprezentovaním prítomnosti alebo absencie proteínov, použitím boolovských rovníc, výpočtová náročnosť simulácie difúzie pomocou deferenciálnych rovníc je eliminovaná.
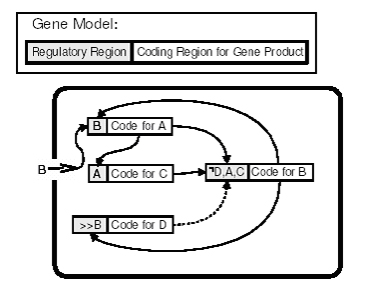
Genetická regulačná sieť
V tomto príklade každý z génov je modelovaný ako regulačný región a kódovací región, ktorý kóduje čiastkový produkt (napr. proteín v prírodnej bunke). Je to jednoduchá sieť (oblý obdĺžnik) vnútri bunky, ktorá ukazuje ako rôzne génové produkty niektorých génov regulujú ostatné gény. Táto sieť opisuje systém, ktorý produkuje niekoľko produktov a potom sa vypne, ak bolo vyrobených dosť produktov B. Symbol „>>“ znamená veľké množstvo. Graf ukazuje, že celá sieť sa stáva aktívnou, ak produkt B vstúpi do bunky z vonkajšieho zdroja. B zapríčiňuje produkciu A, ktoré zase zapríčiňuje produkciu C iným génom. A a C, bez D prítomného v bunke zapríčiňujú výrobu väčšieho množstva B, ktoré zase spätne ovplyvňuje výrobu A, čo ďalej zosilňuje cyklus. Ak však je prítomné veľké množstvo B, vyrobí sa D, čím sa zastaví výroba B a takto sa ukončí spätnoväzobný cyklus . V modeli UE môžu gény napríklad zapríčiňovať rast axónov alebo redukovať neurónový prah.
Viacero nízkoúrovňových biologických procesov používa proteíny produkované operónmi. Napríklad môže byť vložené zariadenie, ktoré zavedie nový proteín na receptor inej bunky. Bunky vyrastajú svoje axóny ak majú špeciálny proteín pre rast axónov v svojej cytoplazme. Axóny sa potom napoja na bunky so správnym cieľovým proteínom. Takto sa nervový systém môže vyvíjať biologicky priateľným spôsobom.
Bunky sa v simulácii delia, ak je prítomný špeciálny deliaci proteín. Po rozdelení majú obidve nástupnícke bunky rovnaký stav, čo vedie k problémom. Ako môže byť symetria vôbec porušená, aby sa mohla uplatniť diferenciácia buniek? Model rieši tento problém prenesením jediného symetriu porušujúceho proteínu po prvom delení bunky, čím umožňuje komunikáciu medzi bunkami.
GRN model odhalil dôležitú výzvu pre UE. (Dellaert a Beer, 1996) ručne naprogramovali genóm pre morfológiu tela (senzory a akčné členy) a pre nervový systém jedinca, vozidla, ktorý sa dokázal vyhýbať prekážkam. Potom boli schopní evolvovať zlepšené verzie jedinca z počiatočného ručne kódovaného genómu.
Nepoddajné prehľadávacie priestory sú jasnou výzvou pre UE systémy vo všeobecnosti. Preto, tak ako gramatické prístupy aj prístupy bunkovej chémie sa musia venovať svojmu biasu a evolvovateľnosti